to the addition of a and � napthylacetate as substrates. Those esterases maintaining activity were regarded Heat stable esterases (HsEST).(33, 34)
International Journal of Scientific & Engineering Research Volume 4, Issue 1, January-2013 1
ISSN 2229-5518
Fitness components of bivoltine breed CSR2 after introgression of multivoltine thermotolerance character of silkworm Bombyx mori L
Gopal Chandra Das a, , Bharat Bhusan Patnaik b and Debjani Nath c*
Bivoltine rearing in tropical regions is highly unpredictable due to fluctuating temperature and humidity conditions. It is therefore extremely imperative to develop a stable bivoltine cocoon crop under adverse environment of the tropices. Improvement of the existing bivoltine lines through introgression of true multivoltine tolerant factors by conventional backcross breeding strategy leading to the development of Near Isogenic Lines (NILs) can bring sustained boon in the sericulture industry. A targeted breeding approach was initiated towards the improvement of productive bivoltine breed ‘CSR2’, developed in South India. In this regard, ‘Nistari’ and ‘Sarupat’ (indigenous multivoltine thermotolerant breeds of West Bengal and Assam respectively) were selected as donor parents for introgression of thermostable factors. Protein profiling of haemolymph indicated the presence of 102-kDa, 89 KDa and 86 KDa polypeptide in the indigenous multivoltines, but absent in developed bivoltine ‘CSR2’. These proteins, as characteristic feature of thermotolerant multivoltine breed, have been successfully integrated in NILs. Esterase isozyme analysis revealed the presence of thermostable esterases (both specific and non-specific) in NILs of
‘CSR2’. In case of a-esterase three bands (Est1,Est2 and Est3) are present in multivoltine parent but in CSR2 Est3 is absent . Heat
stable Est1 and Est2 have successfully integrated in all NILs of which Est2 is heat sensitive in CSR2. This marks the introgression of multivoltine thermostable factors into the improved NILs of bivoltine Silkworm breed ‘CSR2’ leading to improved survivability measured in terms pupation percentage that will help generate bivoltine seed cocoons in this zone for sustaining the silk industry.
The susceptibility of organisms inhabiting tropical climate is likely to differ from those from temperate climate. This aspect has direct relation with the production of silk as temperature and humidity plays major role on growth and productivity of silkworm. The native multivoltine races of Bombyx mori (Sarupat and Nistari strains) exhibit much more tolerance to high temperatures, often the field temperatures touching 40°C or more in summer, as against the exotic bivoltine races (CSR2) of temperate origin (1,2,3).
a Research Extension Centre ( Sub-Unit), Kokrajhar, BTC, Assam
bCentral Sericultural Research & Training Institute, Berhampore-
742101, Murshidabad, West Bengal
c Department of Zoology, University of Kalyani, Kalyani, Nadia West
Bengal, India
* Corresponding author
Tel no. 91-33-24109031 Fax no. 91-33-25828282
E mail address: nath_debjani@yahoo.co.in
The latter strains are better yielder of silk fiber in terms of quality and quantity but less popular with the farmers due to their higher susceptibility to fluctuations in temperature.
In tropical climate sericulture being mostly
multivoltine breed oriented is restricted to rearing of bivoltine for preparation of hybrids (as male component). Due to poor adaptability of these breeds in the fluctuating agro-climatic condition of the tropics, it is therefore imperative to utilize the recurring back cross breeding scheme coupled with selection of growth rate pattern and economic traits in relation the target / desired traits.
( 4) developed two bivoltine breeds MG511 and MG512 of Silkworm for higher viability and silk productivity by conventional back cross breeding. Two promising breeds namely D6 (P) N and SK3C were developed from
IJSER © 2013 http://www.ijser.org
International Journal of Scientific & Engineering Research Volume 4, Issue 1, January-2013 2
ISSN 2229-5518
combination of D6 (P) x Nistari and SK3 x Cambodge by back cross breeding approach (5). Similarly Gangwar et al. (2009) (6) synthesized a promising bivoltine breed UP1 of the Silkworm Bombyx mori L. for the tropical climate of India.
A number of protein substances are usually accumulated in the hemolymph Hemolymph is very important for the metamorphosis of insects including silkworm (7,8) Omana and Gopinatha (1995) (9) have shown a change in protein profile in tissues and haemolymph following heat shock of B mori in vivo.
Understanding the genetic constitution of an
individual in the population of races and allelic variations through isozyme studies is known to reflect the differential catalytic ability of allelic genes and their significant role in the adaptive strategy of the genotypes (10,11) . Among the various known isozymes, esterases have been studied extensively since they are the group of enzyme involved in the metabolic processes of fat body of silkworm involved in gonadal maturation, maintenance of cell viability, metabolic activities of silk gland and defense functions (12,13,14,15,16,17). Isozyme polymorphism has been less studied in Bombyx mori L (18,19), than genetically determined polymorphism in different enzymes (14,
20,21.22,23, 24, 25). The thermo tolerance is positively related to the activity of a heat-stable esterase (Hs EST) analyzed in larval body or midgut. This enzyme activity could be adopted as an indicator for the silkworm thermo tolerance. Wu and Hou (1993) (26) showed that an E5 band in midgut preparations is predominant and is thus related to thermo tolerance and the activity of Hs EST, especially E5, increased with rear selection for 7 generations, indicating that this enzyme seems to be induced by rearing the silkworms under high temperatures.
The present breeding programme was initiated
towards development of near-isogenic lines of promising bivoltine breed CSR2 having high shell weight, SR%, filament length by introgressing the target gene of thermo- tolerance from Nistari (Indigenous breed of West Bengal) and Sarupat (Indigenous breed of Assam). This would help in developing a sustainable breed depending on hemolymph protein profile and isozymic variation in adverse fluctuating high temperature and humid regions of tropics.
‘CSR2’, a (oval shaped and white cocooned) productive
bivoltine breed of Silkworm Bombyx mori L with high qualitative and quantitative traits was selected as recurrent parent. Robust native multivoltine breeds viz., Nistari (elliptical and yellow cocoons) and Sarupat (elliptical and white cocoons) were selected as donor parents carrying target gene(s) for thermotolerance.
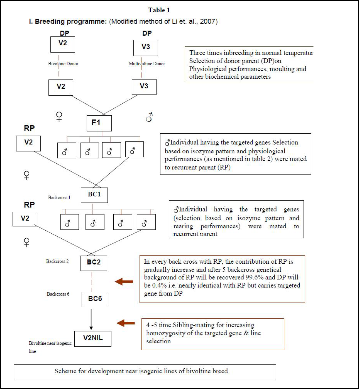
At the initiation of the breeding plan (Table1), crosses were made between female recurrent bivoltine
‘CSR2’ and male donor multivoltine breeds ‘Nistari’ or
‘Sarupat’ separately. Each F1 progeny was backcrossed with recurrent bivoltine ‘CSR2’ (BC1, BC2, BC3…) towards regaining productive and qualitative traits. After six backcross sibling-mating and batch selection was performed on the basis of improved survival rate (measured in terms of effective rearing rate and percentage of pupation) and the presence of thermostable factors through protein profiling and esterase zymographic studies. In addition, during course of breeding experiments other qualitative and quantitative traits (like hibernating, fecundity, larval duration, shell weight, shell%, FL, silk percentage.) were also considered. From F1 to the development of NILs, batch rearing was conducted under ambient temperature (25 ± 3oC) and humidity ranging from
70-90% ( 27). Data collected were subjected to ANOVA
analysis.
Larval haemolymph samples (Vth instar, 5th day) were collected by puncturing proleg of the sixth abdominal segment and drained into eppendorf tube kept in crushed ice with sufficient quantity of phenylthiourea and centrifuged subsequently at 6000 rpm for 10 minutes to
remove haemocytes and other tissue debris (28).
Polyacrylamide gel electrophoresis under denaturing conditions (SDS-PAGE) was performed using
5% stacking and 10% resolving gel (29) with some modifications. Amount of protein was estimated using the method of Bradford, (1976) (30). Electrophoresis was performed for 6hr at 70V for stacking gel and 150V for resolving gel.The gel was stained with 0.2% coomassie brilliant blue R 250. Molecular weight of sample bands were drawn by using reference molecular weight standard(Bangalore Geni India) and compiled utilizing Gel documentation software(UVP GDS 7600,USA)
IJSER © 2013 http://www.ijser.org
International Journal of Scientific & Engineering Research Volume 4, Issue 1, January-2013 3
ISSN 2229-5518
to the addition of a and � napthylacetate as substrates. Those esterases maintaining activity were regarded Heat stable esterases (HsEST).(33, 34)
The back cross lines developed as BC6 NIL (S) & BC6 NIL (N) from the combination of recurring bivoltine parent CSR2 with donor multivoltines, Sarupat and Nistari (Table 1) respectively recorded promising yield performance during unfavorable seasons. The pupation % (Figure1) and ERR % recorded was higher in BC6 NIL (S)
compared to back cross line developed with Nistari, but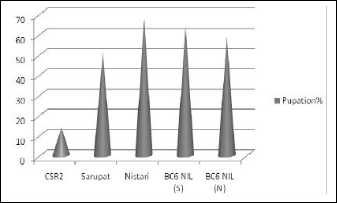
Equal amount of each samples were electrophoresed under non-denaturing conditions on a 8% polyacrylamide gel following Pharmacia Laboratory techniques using dual vertical gel electrophoresis system attached to Thermo-controlled water bath (Pharmacia LKB
– MultiTemp II). Polyacrylamide gel electrophoresis
(PAGE) was carried out under 110 V constant voltage for 3-
4 hr at 4oC until the tracing dye reached the bottom of the gel.
Figure 1. Comparative performance of pupation (%) in the BC6 lines developed so far with donor and recipient
fecundity was found higher in BC6 NIL (N). The performances of BC6 lines (Table 2) in relation to other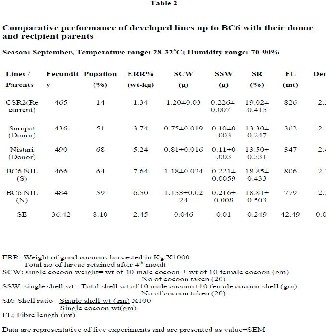
Esterase isozyme patterns were studied by staining the gels in the presence of a and � napthylacetate as substrates by modified method of Johnson and Deniston (1964) and Simms (1965) (31,32). Following electrophoresis, the gels were immersed in 0.5 M Boric acid (pH 4.1) for 30 min at 4oC. Gels were then rinsed rapidly in two changes of
ice-cold distilled water and placed separately in trays with substrate of 2% a or � napthylacetate in acetone, 40 mg of Fast Blue BB salt and 100 ml of 0.2 M sodium phosphate buffer A (pH 9.2) and B (pH 4.3). Gels were incubated under dark conditions at 30oC for 4-8 hr. Gel documentation was performed with the GDS-7600-UVP
white/UV transilluminator.
Normal gels of haemolymph samples after electrophoresis
were incubated at 70 ± 2oC in a water bath for 10 min prior
IJSER © 2013 http://www.ijser.org
International Journal of Scientific & Engineering Research Volume 4, Issue 1, January-2013 4
ISSN 2229-5518
significant yield parameters viz., SCW, SSW and SR% were at par with their respective parents. Post -cocoon analysis indicated a decline of 2.25% and 5.70% in FL for BC6 NIL (S) and BC6 NIL (N) respectively.
Results revealed highly significant (P < 0.01) increase of survivability in the developed lines so far in comparison to the recurrent parent, which is reflected in higher pupation percentage. About four-fold increase of rate of survival in BC6 NIL (S) and BC6 NIL (N) have been observed in comparison to the recurrent parent CSR2. This indicates the successful introgression of survival genes into the near isogenic lines of CSR2 without compromising on the quality parameters.
One-dimensional gel electrophoresis of the haemolymph samples of recurrent back cross lines in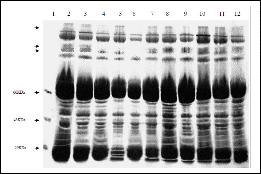
2A
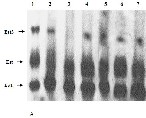
Figure2 A SDS Gel electrophoretic protein profile of donor , recipient parents and developed breeds. Lane 1: Marker (Bangalore Genei); Lane
2: Nistari (donor parent), Lane 3: Sarupat (donor parent), Lane 6: CSR2 (recurrent parent), Lane 4: BC3NIL (S), Lane 5: BC4 NIL (S), lane 7: BC5
NIL (S), lane 8: BC6 NIL (S), Lane 9: BC6 NIL (S)2, Lane 10: BC4 NIL (N), Lane 11: BC5 NIL (N), lane 12 : BC6 NIL (N).
2B Scanning of SDS gel protein profile A Nistari; B Sarupat; C CSR2;
D BC4 NIL (S); E BC5 NIL(S); F BC6NIL(S); G BC4NIL(N); H BC5NIL(N); I BC6NIL(N)
comparison to their respective parents was conducted to analyze the introgression of gene through the expression of targeted polypeptides. Protein profiling. Showed (Fig 2A, B) major proteins in parent Nistari are of 102 KDa,
98KDa,94KDa, 89KDA, 86KDa, proteins of molecular wt in
between 66KDa and 54 KDa, 52 KDa ,46KDa, 41KDa , major proteins of 39KDa, 35KDa, 30KDa, 29KDa and 26KDa ; in Sarupat most proteins are of same mol wt except comparatively less amount of 42KDa ,35 K Da and 30 K Da protein. It is striking point to note the absence of three proteins like 102KDA, 89KDa and 86KDa and less amount of proteins at the level of 52 K Da and 30 KDa. in recurrent parent CSR(2) in comparison to donors Nistari and Sarupat. The presence of the same proteins like 102 KDA, 89 KDa and 86 KDa in the BC6 lines of CSR2 of both donor Sarupat and Nistari and increased amount of protein at the level of
30KDa are evidence of genetic introgression during
crossing .
2B lines of CSR2:
8% native PAGE with a and � naphthyl acetate as
B substrate study in haemolymph protein expression of
C
specific and non-specific protein in (all backcross lines up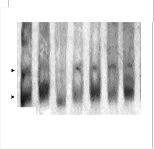
to BC6)NIL of CSR2and their parents shows polymorphism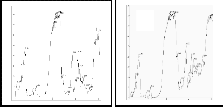

D E F
1 2 3 4 5 6 7
B

G H I
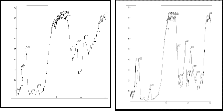
Figure 3. A.Vertical 8% Page of a –esterase enzyme of haemolymph of B mori parents and developed breeds.Bands Est
1and 2 were specific and 3 was nonspecific using 2% naphthylacetate as substrate. Specific and nonspecific a-esterase isozyme bands were identified by the absence and presence in the PAGE of � -esterase isozyme pattern of the same sample using �-naphthylacatate as a substrate.B. heat stable a-esterase banding pattern . these bands were observed only when normal gel of haemolymph sample was incubated at 70 ± 2oC in a water:
IJSER © 2013 http://www.ijser.org
International Journal of Scientific & Engineering Research Volume 4, Issue 1, January-2013 5
ISSN 2229-5518
bath for 10 min prior to the addition of a napthylacetate as substrates Lane 1: Nistari; Lane 2: Sarupat; Lane 3: CSR2; Lane 4: BC5NIL(N); Lane 5: BC6NIL(N); Lane 6: BC5NIL(S); Lane 7: BC6NIL(S)
in nonspecific a-esterase and found 3 bands (Est-1, 2 and
3,) (Fig 3) in haemolymph of whereas CSR2 has two bands,
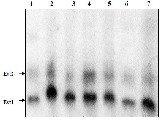
.No polymorphism found in �-esterase isozyme pattern having two bands (Est-1 & 2) in haemolymph of silkworm Bombyx mori L.(Fig 4). The polymorphisms of nonspecific esterase, their genetic variability in inter breed of Bombyx mori L is critical during selection of improve strains (35).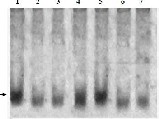
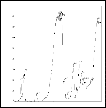
A B
Figure 4 A Vertical 8% Page of � –esterase enzyme of haemolymph of B mori parents and developed breeds. Bands Est 1 and2 were specific using 2% naphthylacetate as substrate. Specific and nonspecific a- esterase isozyme bands were identified by the absence and presence in the PAGE of a � -esterase isozyme pattern of the same sample using a- naphthylacatate as a substrate. B. heat stable � -esterase banding pattern . These bands were observed only when normal gel of haemolymph proteins was incubated at 70 ± 2oC in a water bath for 10 min prior to the addition of � -napthylacetate as substrates Lane 1: Nistari; Lane 2: Sarupat; Lane 3: CSR2; Lane 4: BC5NIL(N); Lane 5: BC6NIL(N); Lane 6: BC5NIL(S); Lane 7: BC6NIL(S)
Zymograms showed that the specific a–esterase band Est 1 and 2 and specific �- esterase bands (Est. 1) which have been successfully integrated predominately withstand the temperature of 70 0C for 10 minutes and thereby reported as thermostable esterase in haemolymph in silkworm Bombyx mori L.
Two basic requirements in breeding – are the presence of genetic variation and exploitation of this variation through selection. The presence of genetic variation in populations, especially the favorable and beneficial characters, is a must for any improvement programme. If all the individuals in a population were identical for a character, variation can be created through hybridization, mutation etc. This is followed by selection of desirable individuals for improvement of the traits. This paves the way to amalgamate more desirable characters into a single breed that may have acquired buffering capacity against the prevailing environmental
impediments. Phenotypic selection may not entirely characterize the variations present due to environmental influence on the expression potential of desired genes, thus genomic and proteomic tools can sufficiently assist the conventional breeders to demarcate the subtle changes in a population.
NIL populations have been extensively used in
genetic studies due to the advantages derived from their homozygosity (36, 3, 37). NILs are considered important material to locate the phenotypic variations between pairs of lines that can be assigned directly to the distinct genomic regions introgressed in another similar genetic background (38). In this research, NIL of CSR2 were being constructed with the aim of utilizing the eventual breed in the tropical climatic conditions of India by introgression of desired thermo tolerant genic variations from indigenous multivoltines viz. Sarupat and Nistari. BC4 NIL (S) and BC4
NIL (N) thus evolved so far depicts improved survivability
without losing the yield efficiency (Table2). Since improvement of the rate of survivality was required in productive bivoltines, back crossing was adopted (39) (Das,
2001) and bi x multi crosses were considered as base
material for breeding. The present study clearly indicated a step closer towards improvement of survivability in developed bivoltine, NIL of CSR2, comparing to their original parents.
Theoretically, after every back cross,
contribution of the recurrent parent improves by 50% and a line is obtained which carries the target gene in its genetic background (40, 41). As the genetic background of NIL is similar with the recurrent parent except the target genes received from donor parent, protein profiling will be different in the region of the target gene leading to unique expression of proteins. Polypeptide profiling (Figure2) clearly signifies the presence of 102kDa , 89KD a and
86KDa bands in BC4 NIL (S) and BC4 NIL (N) but not in bivoltine CSR2. Significant quantitative integration of polypeptides corresponding to 52 and 30kDa were also found in NILs suggesting their role as adaptor proteins. The expression of these proteins may be related to the heat shock tolerance of Nistari and Sarupat which has also been supported the development of heat tolerance of near isogenic lines of the multivoltine breeds. Generally, the heat shock response depends on the magnitude of temperature elevation and duration of exposure and is relative to the environmental temperature at which the organism normally survives (42,43, 9) In addition, expression of HSPs in different tissues varied depending on the stage of
development, the temperature, and/or at which stage exposure was performed (44, 45)). Manjunatha et.al.
(2010)(46) indicated that different sets of HSPs were being
IJSER © 2013 http://www.ijser.org
International Journal of Scientific & Engineering Research Volume 4, Issue 1, January-2013 6
ISSN 2229-5518
expressed at various heat shock temperatures, in different breeds of B. mori of which 90 and 84 kDa HSPs were ubiquitous. Notably, concentration of HSPs and their distribution to specific sub-cellular sites is an important factor in acquisition of thermotolerance (47). So there is a need to develop the screening procedure for identifying levels of acquired thermo -tolerance led to the evaluation of different cellular constituents that might be used as an In
vivo indicator of heat injury or cell viability and further
studies are needed to understand whether 102kDa , 89KD a and 86KDa polypeptides or over expressed 52 K Da or 32
KDa polypeptides act as only HSPs or as phenotypic
expression of new protein background supporting thermotolerance in changing genetic environment of the NILs.
Tansley and Rick (1980)(48) exploited differences in isozymic allele between the donor and recurring parent.
Liu et.al.(1984)(34) tried to develop thermotolerant silkworm strains after induction of higher temperature and
observed the appearance of the heat stable esterases in these strains. We observed that two heat stable a–esterase bands in donor parents Nistari and Sarupat and their near isogenic lines whereas in CSR2 one band was observed both in bivoltine and multivoltine breeds(Fig 4), has any relation with thermotolerance . If so this could be also utilized as a biochemical marker to screen thermotolerant breeds or strains of Bombyx mori
. The authors are deeply indebted to Vice chancellor
,University of Kalyani and Director, Central Sericulture Research and Training Institute, Berhampur, Murshidabad for their interest and support in this research. There is no conflict of interest regarding the primary interest as well as secondary interest of authors on the study in the mentioned field of research and publication
.
1. Krishnaswamy S, Narasimhanna M N,
Suryanaryan S K and Kumararaj S 1977 Manual on sericulture Vol. 2 Silkworm rearing (Mysore: Publ. Central Silk Board)
2. Pershad G D, Datta R K, Vijayakumar H V,
Bhargava S K and Jolly M S 1986 Performance of
some multivoltine races of Bombyx mori L;
Sericologia 26 295–301
3. Suresh Kumar N., 2Harjeet Singh, 1Saha A. K. and
1Bindroo B. B. (2011) Development of Bivoltine double hybrid of the silkworm, Bombyx Mori L. tolerant to high temperature and high humidity conditions of the tropics.Universal Journal of Environmental Research and Technology Volume
1, Issue 4: 423-434
4. Raju, P.J., Krishnamurthy, N.B., 1993. Breeding of two bivoltines, MG 511 and MG 512 of Silkworm, Bombyx mori L. for higher viability and silk productivity. Sericologia 33, 577-587.
5. Moorthy, S.M., Das, S.K., Rao, P.R.T., Raje U, S., Sarkar, A., 2007. Evaluation and selection of potential parents based on selection indices and isozyme variability in silkworm, Bombyx mori L. Int. J. Indust. Entomol. 14(1), 1-7.
6. Gangwar, S.K., Jaiswal, K., Dwivedi, P., Gupta, V.,
2009. Synthesis of promising bivoltine breed UP1 of the silkworm (Bombyx mori L.) for Uttar Pradesh. ARPN J. Agril. Biol. Sci. 4(4), 8-12.
7. Taniai K and Yamakawa M1998 Expression of anti
bacterila protein gene in ligated larvae of silkworm
Appl Ent Zool 33: 179-183
8. Kajiwara H, Imamaki A; Nakamura M; Mita R; Xia QG; Ishizaka M.(2009) Proteome analysis of silkworm. 1.Fat body J Electrophoresis.53: 19-26
9. Omana J and Gopinathan K P 1995 Heat shock
response in mulberry silkworm races with different thermotolerances J. Biosci., 20, p 499–513.
10. Parkash R; Yadav J P and Vashisht M. 1992.
Allozymic variation at ADH locus in some
Drosophila species. Perspectives in Cytologyand
Genetics 8: 495-502.
IJSER © 2013 http://www.ijser.org
International Journal of Scientific & Engineering Research Volume 4, Issue 1, January-2013 7
ISSN 2229-5518
11. Pushpalatha N and Vijayan V A 1999. Isozyme profiles in relation to ecological status in two Japanese encephalitis vectors. Culex vishnui and Culex Fuscocephala Diptera: Culicidae) Entomon 24:
297-306.
12. Yoshitake N and Akiyama M 1965. Genetic aspect on the esterase activities of the egg in the silkworm BomlJyx mori L (in Japanese) lpn. Seri cult. Sci. 34:
327-332.
13. Yoshitake N and Eguchi M 1965. Distribution of blood esterase types in various strains of the silkworm BomlJyx mori L (in Japanse) lpn.Sericult. Sci. 34: 95-98.
14. Eguchi M ; Yoshitake N and Kai H. 1965. Types
and inheritance of the blood esterase in the silkworm, Bombyx mori L. J Genet. 40: 15-19.
15. Eguchi M and Yoshitake N. 1967. Interrelation of
non-specific esterase among various tissues in the silkworm, Bombyx mori L. lpn. J Seri cult. 36: 193-
198.
16. Bhattacharyya A ; Sarkar S K; Ganguly S and Basu S K 1990. Studies on the isozymes patterns of G-6- PD and LDH in male pupa of Eri-silkworm, Philosamia ricini. Science and Culture 56: 131-132
17. Somasundaram P; Ashok Kumar K; Thangavelu K;
Kar PK and Sinha R K 2004 Preliminary Study on Isozyme Variation in Silkworm Germplasm of Bombyx mori (L.) and its Implication for Conservation Central Sericultural Germplasm Resources PertanikaJ. Trop. Agric. Sci. 27(2): 163 -
171
18. Shabalina A., 1990. Esterase genetic polymorphism in haemolymph of larvae Bombyx mori L. Compets rendus de I’Academie bulgare des sciences, 43: 105-108.
19. Stoikova T., Popov P., Dimitrov B., 2003.
Electrophoretic analysis of non specific haemolymph esterases during silkworm (Bombyx mori L) ontogenesis. Sericologia 43: 153-162.
20. Eguchi M., Takahama Y., Ikeda M., Horii S., 1988.
A novel variant of acid phosphatase isozyme from haemolymph of silkworm, Bombyx mori L. Japan Journal of Genetics, 63: 149-157
21. Eguchi M., 1995. Alkaline phosphatase isozymes in
insects and comparison with mammalian enzyme. Comparative Biochemistry and Physiology, 111B:
151-162.
22. Hara, W., Pujii H., Sakaguchi B., 1984. Activity and polymorphism of amylase in gut contents in different lines of Bombyx mori L. Journal of sericultural Science of Japan, 53: 496-500.
23. Egorova T., Naletova E., Nasirillaev Y. 1985.
Polymorphic system of silkworm haemolymph esterases as a criterion to make programs for parental specimens crossing. Biochemistry of insects –Moskow 54-62.
24. Takeda S., Azuma M., Eguchi M., 1990. Alkaline
phosphatase isozymes of the silkworm midgut: polymorphism in the enzyme activity, antigen and electroforetogram. Journal of sericultural Science of Japan, 59: 127-134.
25. Yamamoto K., Yakiyama M., Pujii H., Kusakabe T.,
Koga K.,Aso Y., Ishioguro M., 2000. Expression of prophenoloxidase m-RNA during hemocyte development. Bioscience, Biotechnology and
Biochemistry, 64: 1197-1202
26. Wu D J ;Hou R F 1993 Relationship between thermotolerance and heat stable esterases in the silk worm Bombyx mori L (Lepidoptera: Bombycidae)Appl entomology Zool 28:371-377
27. Krishnaswami, S., 1978. New technology of
silkworm rearing. Bulletin No.2, Central Sericultural Research and Training Institute (Mysore), Central Silk Board, India, pp. 1-23.
28. Gupta, A.P., 1979. Haemocyte Types: Their
Structure, Synonymies, Interrelationships and Taxonomic Significance. In: Insect Haemocytes, Gupta, A.P. (Ed.). Cambridge University Press, Cambridge, pp: 85-127
29. Laemmli, U.K., 1970. Cleavage of structural
proteins during the assembly of the head of bacteriophage T4. Nature 227, 680-685.
30. Bradford, M.A., 1976. Rapid and sensitive method
for the quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72, 248-254.
31. Johnson, F.M., Denniston, C., (1964). Genetic
variation of alcohol dehydrogenase in Drosophila melanogaster. Nature, 204: 906-907
32. Simms M (1964) Methods for detection of
enzymatic activity after electrophoresis in polyacrylamide gel in Drosophila sp. Nature
:207,757-758
33. Cochrane B J (1976) Heat stability variants of esterase 6 in Drosophila melanogaster Nature, 263:
131-132
34. Liu Z G ; Kabayashi M and Yoshitake N (1984) Genetical studies on the heat stable expression in midgut of the silkworm, Bombyx. Mori J steric Sc Jpn (in Japanese)
35. Patnaik A., Datta R. K., 1995. Amylase – its
genetics and prospects as a marker in silkworm breeding. Indian Journal of sericulture, 34:82 – 89.
IJSER © 2013 http://www.ijser.org
International Journal of Scientific & Engineering Research Volume 4, Issue 1, January-2013 8
ISSN 2229-5518
36. Chattopadhyay, G.K., Sengupta, A.K., Verma, A.K., Sen, S.K., Saratchandra, B., 2001. Transgression of shell weight- - A multigenic trait through development of congenic breed in tropical silkworm, Bombyx mori L. Sericologia 41(1), 33-42.
37. Li, M., Hou, C., Zhao, Y., Xu, A., Guo, X., Huang,
Y., 2007. Detection of homozygosity in near isogenic lines of non-susceptible to Zhenjiang strain of densonucleosis virus in silkworm. Afric. J. Biotechnol. 6(14), 1629-1633.
38. Nadeau, J.H., Singer, J.B., Matin, A., Lander, E.S.,
2000. Analyzing complex genetic traits with chromosome substitution strains. Nat. Genet. 24,
221-225.
39. Das, S.K., 2001. Techniques of breeding for evolving improved breeds of bivoltine mulberry silkworm, Bombyx mori L. Persp. Cytol. Genet. 10,
129-134.
40. Chattopadhyay, G.K., Verma, A.K., Sengupta, A.K., Das, S.K., Raje Urs, S., 2004. a- and �- amylase isozyme expresser native proteins in tropical silkworm, Bombyx mori L. Int. J. Indust. Entomol. 8(2), 189-194.
41. Sreekumar, S., Ashwath, S.K, Nirmal Kumar, S., Kamble, C.K., 2008. Molecular Markers as selection tools foe gene tagged breeding in the mulberry silkworm, Bombyx mori L. Breeders meet, CSR&TI, Mysore, pp 36-40.
42. Nakayama S; Suzuki A Fujii S, Yamamoto R 2001
Sugar detection with fluorescein isothiocyanate- labeled lectins in hemolymph proteins of silk worm Bombyx mori (Lepidopteran,Bombycidae) Appl EntomolZool 36:439-442
43. Nath B B and Lakhotia S C 1989 Heat shock
response in tropical chironomus: seasonal variation in response and the effect of developmental stage and tissue type on heat shock protein synthesis.Genome 32 676–686
44. Joy o, gopinathan K P (1995) Heat shock response
in mulberry silkworm races with different thermo tolerance J Bioscience 20:499-513
45. Vasudha B. C; Sosalegowda A H and Manjunath
H B (2006) Impact of heat shock on heat shoch protein expression, biological and commertial traits of Bombyx mori. Insect Science 13: 243
46. Manjunatha H B; Rajesh R K and Aparna H S
(2010) Silkworm thermal biology: A review of heat shock response, heat shock proteins and heat acclimation in the domesticated silkworm Bombyx mori. J Insect Science 10: 204
47. Kampinga H H ;Thermotolerance in mammalian
protein denaturation and aggregation and stress protein (1993). J Cell Sci 104: 11-17
48. Tansley, S.D., Rick, C.M., 1980. Isozyme gene
linkage map of the tomato: application in genetics
and breeding. Theor. Appl. Genet. 57, 161-170
IJSER © 2013 http://www.ijser.org