International Journal of Scientific & Engineering Research, Volume 5, Issue 2, February-2014 209
ISSN 2229-5518
Effects of Competition from the Invasive Cordgrass Spartina densiflora on Native Atriplex portulacoides
Ahmed M. Abbas, Enrique Figueroa, Alfonso de Cires, Alfredo E. Rubio-Casal, Jesús M. Castillo
Abstract— Invasion by the South American cordgrass Spartina densiflora Brongn. in European salt marshes is causing concern about potential impacts to native plant communities., S. densiflora colonization of mid-marsh habitats may be slowed by native Atriplex portulacoides L., but the mechanism behind this negative interaction is not known. The aim of this work was to study the establishment and growth response of S. densiflora seedlings grow- ing in a competitive environment with mature A. portulacoides plants in controlled greenhouse conditions. W ith this aim we measured establishment, growth, foliar nutrients, and photosynthetic pigments of S. densiflora plants grown with and without A. portulacoides. Results showed that S. densiflora seedlings readily established in mature stands of A. portulacoides. Every Spartina clump growing with Atriplex survived, producing fewer tillers with simi- lar heights than when growing without competition, which was reflected on lower above-ground biomass. These results indicated that S. densiflora was affected at the above-ground level by the interspecific competition with Atriplex, but the invasive cordgrass was able to keep similar below-ground bio- mass with and without competition, resulting in a decrease of below-ground biomass of Atriplex when competing with Spartina. Our results in green- house controlled conditions were in agreement with our field observations where S. densiflora plants have been observed growing within A. portu- lacoides monospecific communities.
Index Terms— Biomass accumulation, Brackish marshes, Competitive interactions, Greenhouse, Invasion, Salt marshes, Shoot production.
1 INTRODUCTION
—————————— ——————————
ntroduced invasive plants to natural areas sometimes threaten local communities and decrease native diversity [1], potentially through competitive mechanisms [2]. In this context, experimental studies to determine the outcome of interspecific competition between invasive and native species are necessary for conserving and restoring native communities
in impacted habitats [3].
Plant distribution in salt marshes is determined by species
tolerance to abiotic factors and by interspecific interactions
such as predation, facilitation and competition [4]. Thus, inter- specific competition is one of the most important processes determining plant distribution in salt marshes [5]. The intro- duction of alien halophytes may alter natural zonation pat- terns in salt marshes [6].
The South American cordgrass Spartina densiflora Brongn. (Poaceae) is invading estuaries in southwest Europe, north- west Africa and the West Coast of North America [7]. In The Gulf of Cádiz (southwest Iberian Peninsula), S. densiflora is colonising middle marsh communities dominated by Atriplex portulacoides L. Atriplex portulacoides L. is a halophyte that colonises low marshes, getting its maximum biomasses at middle marshes [8]and producing large quantities of seeds all around the coasts of Europe, North Africa and southwest Asia [9].
————————————————
• Ahmed M. Abbas lecturer in botany department-South valley university- Egypt. He obtained his PhD degree in Plant ecology from Sevilla Universi- ty – Spain with a Distinction in 2012. PH- +2(0)1124094908 E-mail: abbas@sci.svu.edu.eg
• Second, third, fourth and fivth author: Associate Professor, Assistant Pro-
fessor, Professor and Assistant Professor, Department of Plant Biology and Ecology, University of Seville, Av. Reina Mercedes, s/n.-41012 Sevilla – Spain. Jesús M. Castillo: manucas@us.es, Enrique Figueroa: Figueroa@us.es, Alfonso de Cires: decires@us.es, Alfredo E. Rubio-Casal: aerubio@us.es
The aim of this work was to study the establishment of S. densiflora seedlings growing together with adult A. portu- lacoides plants in controlled greenhouse conditions. With this aim we recorded establishment, growth, foliar nutrients and photosynthetic pigments of S. densiflora plants coming from seeds in response to inter-specific competition with A. portu- lacoides and in the absence of competition. Experimental ap- proaches working in controlled conditions have been used frequently to study competition between invasive and native species [10]. In view of our field observations, we hypothe- sized that S. densiflora would be able to establish and compete successfully with A. portulacoides adult plants.
.
2 MATERIALS AND METHODS
2.1 Plant Collection
Seeds of S. densiflora were collected in April 2008 from multi- ple mature individuals in a well-drained intertidal lagoon at the Odiel Marshes (southwest Iberian Peninsula; 37º 08´ - 37º
20´ N, 6º 45´ - 7º 02´ W) [11].
2.2 Experimental Design
The experiment was initiated in May 2008 and conducted over two years in the greenhouse facility of the University of Se- ville, Spain. S. densiflora seeds were planted one cm deep in peat soil in plastic pots (7.5 cm diameter and 12 cm height; volume of 1.85 l) and exposed to two competition treatments (n = 6 pots per treatment): (1) one seed of Spartina per pot (control treatment), and; (2) one seed of S. densiflora germinat- ing in a pot colonised previously by one adult plant of A. por- tulacoides (1 year old), seeds planted in the middle of each pot.
IJSER © 2014
http://www.ijser.org
International Journal of Scientific & Engineering Research, Volume 5, Issue 2, February-2014 210
ISSN 2229-5518
Six adult plants of A. portulacoides were also grown without competition. Plants were maintained at ambient light in the greenhouse and watered with salt water (20 mg l-1), keeping the base of the pots permanently flooded to a height of 3 cm using pools. Mean monthly air temperature during the exper- iment in the greenhouse was 24 ± 0 ºC, varying between 17 ± 0
ºC and 33 ± 0 ºC. Mean monthly air relative humidity was 60 ±
3%, varying between 45 ± 1% and 73 ± 0%.
2.3 Shoot Production, Height and Biomass
The number of S. densiflora live seedlings and their shoots were counted periodically from the beginning of the experiment for every clump in each pot (n = 6). S. densiflora shoot height was measured from the base of the shoot to the tip of the longest leaf (n = 6 shoots, one shoot per pot).
At the end of the experiment (June 2010), all plants were
carefully excavated from the soil and roots were washed to
remove soil particles. Roots and shoots were separated and above- and below-ground biomass (AGB and BGB, respectively) were recorded for all plants (n = 6 plants per treatment). Biomass was determined after drying plant material at 80 ºC for 48 hours.
2.4 Leaf Nitrogen and Carbon Content
Total leaf carbon (C) and nitrogen (N) content were deter-
ples was applied to explore intraspecific differences in plant traits between the two competition treatments. Variances were calculated as the standard error of the mean. All tests were evaluated at a significance level of 0.05.
3 RESULTS
3.1 Seedling Survivorship, and Tiller Number and
Height
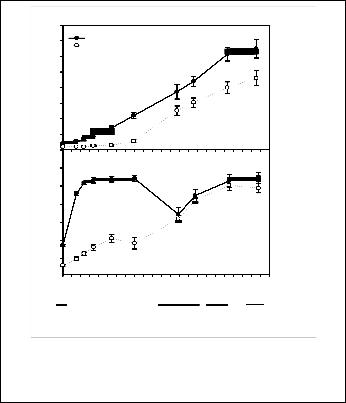
Every S. densiflora clump survived with and without A. portulacoides competition. Competition with adult plants of Atriplex during 26 months decreased the number of live tillers of S. densiflora clumps. Thus, average tiller number for S. densiflora clumps growing alone was 33 ± 3 tillers clump-1 at the end of the experiment and 23 ± 3 tillers clump-1 for clumps growing with A. portulacoides (t-test, t = 2.37, P < 0.05) (Fig. 1a). S. densiflora tillers growing without interspecific competition were taller than those competing with A. portulacoides during the first year, but this difference disappeared since them showing similar maximum tiller height at the end of the experiment (110 ± 5 cm without competition and 98 ± 5 cm with competition; t-test, t = 1.80, P > 0.05) (Fig. 1b).
40
mined in May 2009 for the two plant species. Two leaves from
each plant were chosen randomly and removed from stems by
hand. The samples were dried to constant weight in an oven at
80 ºC for 48 hours, pulverized using a grinder (Cyclotec, Foss Tecator AB, Höganäs, Sweden) and filtered using an 80-μm sieve. Total C and N content of filtered material was deter- mined as percentage of total dry matter for undigested sam-
ples using an elemental analyzer (Leco CHNS-932, Spain). The value for each sample corresponded to the mean of three rep- licated measurements.
2.5 Photosynthetic Pigments
In October 2009, four leaves per treatment were chosen at random from different seedlings to determine leaf photosynthetic pigment content, chlorophyll a (Chl a), chlorophyll b (Chl b) and carotenoids (carotenes and xanthophylls, Cx + c). Pigments wereusing 0.1 g of fresh material in 5 ml of 80% aqueous acetone (n = 4). After filtering,
0.5 ml of the suspension was diluted with a further 2 ml of acetone and Chl a, Chl b and Cx + c concentrations were
35
30
25
20
15
10
5
140
120
100
80
60
40
20
Control
+ A. portulacoides
2008 2009 2010
(a)
(b)
determined with a Hitachi U–2001 spectrophotometer (Hitachi
Ltd., Tokyo, Japan) using three wavelengths (663.2, 646.8 and
470.0 nm). Concentrations of pigments (mg g–1 DW) were
obtained through calculation [12].
2.6 Statistical Analysis
Analyses were conducted using SPSS release 12.0 (SPSS Inc., Chicago, IL). Data were tested for normality with the Kolmo- gorov-Smirnov test and for homogeneity of variance with the Levene test (P > 0.05). A student’s t-test for independent sam-
Fig. 1. Temporal variation in the total number of tillers (a) and maxi- mum tiller height (cm) (b) for Spartina densiflora clumps with and without (control) competition with Atriplex portulacoides.
3.2 Leaf carbon and nitrogen and photosynthetic pigments content
Leaf C content was similar in both competition treatments for both species (Spartina: ~ 44 % C; Atriplex: ~ 38 % C; t-test, P >
IJSER © 2014 http://www.ijser.org
International Journal of Scientific & Engineering Research, Volume 5, Issue 2, February-2014 211
ISSN 2229-5518
TABLE 1

CARBON AND NITROGEN CONTENTS, C:N RATIO, LEAF PHOTOSYNTHETIC PIGMENTS CONCENTRATIONS (CHLOROPHYLL A, CHLO- ROPHYLL B, CAROTENOIDS (CX+C) AND CHLOROPHYLLS : CAROTENOIDS RATIO), AND ABOVE- AND BELOW-GROUND BIOMASS (AGB AND BGB, RESPECTIVELY) FOR SPARTINA DENSIFLORA SEEDLINGS AND ATRIPLEX PORTULACOIDES ADULT PLANTS IN TWO DIFFERENT COMPETITION TREATMENTS (W ITH AND WITHOUT INTERSPECIFIC COMPETITION). DIFFERENT LETTERS INDICATE SIGNIF- ICANT INTRASPECIFIC DIFFERENCE BETW EEN TREATMENTS FOR THE SAME SPECIES (T-TEST, P < 0.05).
Spartina densiflora Atriplex portulacoides
| Without compe- tition | Interspecific com- petition | Without compe- tition | Interspecific compe- tition |
Nutrient content (%) | | | | |
% C | 44.36 ± 0.18a | 44.71 ± 0.16a | 38.04 ± 0.25a | 38.00 ± 0.61a |
% N | 0.75 ± 0.01a | 0.94 ± 0.03a | 1.38 ± 0.02a | 1.08 ± 0.08a |
C/N | 59.03 ± 2.04a | 48.60 ± 2.80b | 27.18 ± 0.50a | 35.82 ± 2.57b |
Photosynthetic pigments | (mg g-1 DW) | | | |
Chl a Chl b | 6.2 ± 1.1a 2.7 ± 0.4a | 5.4 ± 0.3a 2.2 ± 0.4a | 3.0 ± 0.2a 1.2 ± 0.1a | 2.9 ± 0.4a 1.1 ± 0.3a |
Cx+c | 2.5 ± 0.2a | 2.6 ± 0.2a | 0.6 ± 0.0a | 0.5 ± 0.1a |
Chl a + b : Cx+c | 3.4 ± 0.3a | 3.6 ± 0.3a | 7.4 ± 0.6a | 11.5 ± 3.3a |
Biomass (g m-2) | | | | |
AGB BGB | 1256 ± 97a 3085 ± 236a | 987 ± 32b 2867 ± 153a | 817 ± 62a 1869 ± 125a | 758 ± 32a 1522 ± 31b |
0.05). Spartina leaf N content was lower for clumps growing alone than for those growing together with Atriplex (t-test, t =
0.108, P > 0.05). A. portulacoides showed similar leaf N content with and without competition (t-test, t = 0.295, P > 0.05)
(Table 1). The highest C : N ratio was recorded for S. densiflora growing alone, which was higher than for the clumps growing with corresponded to A. portulacoides adult plants growing alone that was lower than with when exposed to competition (t-test, t = -3.222, P < 0.05) (Table 1).
S. densiflora and A. portulacoides showed similar Chl a, Chl b and Cx+c contents and similar Chl a + b: Cx+c ratio for both treatments (t-test, P > 0.05) (Table 1). Atriplex (t-test, t = 3.00, P
< 0.05). The lowest C: N ratio ratio for both treatments (t-test, P > 0.05) (Table 1).
3.3 Biomass
At the end of the experiment, Spartina seedlings growing alone developed higher AGB than when competing with Atriplex (t- test, t = 2.64, P < 0.05), with similar BGB (t-test, t = 0.778, P >
0.05). AGB of Atriplex was not affected by interspecific compe- tition (t-test, t = 0.841, P > 0.05), while its BGB was lower when growing together with Spartina (t-test, t = 2.67, P < 0.05). Spartina clumps accumulated more AGB and BGB than Atri- plex in both treatments (t-test, P < 0.05) (Table 1).
4 DISCUSSION
Every Spartina clump growing with Atriplex survived, produc- ing fewer tillers with similar heights than when growing without competition, which was reflected on lower AGB ac- cumulation. The higher growth of Spartina without competi- tion coincided with a lower leaf N content than when compet- ing with Atriplex (without affecting photosynthetic pigments concentrations), which was probably related with the dilution effect related with faster growth rates [13].These results indi- cated that S. densiflora was affected at the above-ground level by the interspecific competition with Atriplex, but the invasive cordgrass was able to keep similar BGB with and without competition, resulting in a decrease of BGB of Atriplex when competing with Spartina. Thus, the roots and rhizomes of A. portulacoides were not able to prevent the development of rhi- zomes and roots of Spartina seedlings. In contrast with our results, previous studies have recorded a limitation in Spartina development in the underground level when competing with other halophytes. [6] pointed out that the invasion of S. densi- flora from seeds seemed to be limited by inter-specific subter- ranean competition with the native Spartina maritima (Curtis) Fernald. In this sense, it has been described that the invasion of S. densiflora at North American marshes is limited by com- petition with native species [14] and that Typha dominguensis
IJSER © 2014 http://www.ijser.org
International Journal of Scientific & Engineering Research, Volume 5, Issue 2, February-2014 212
ISSN 2229-5518
Pers. and Phragmites australis (Cav.) Trin. ex Steud., limited the subterranean development of S. densiflora . However A. portu- lacoides has been described as well-adapted to competitive environments, being the distribution and frequency of species of its communities mainly conditioned by interspecific compe- tition, [15]. [16], recorded that it was being displaced from some European marshes by the invasive grass Elymus atheri- cus.Our results in greenhouse controlled conditions corrobo- rated our hypothesis based on field observations since the presence of adult plants of Atriplex did not prevented the es- tablishment and growth of S. densiflora that competed success- fully with the native halophyte as observed in the field.
5 CONCLUSION
In this report, the invasive cordgrass S. densiflora is colonizing middle salt marshes in the southwest Iberian Peninsula where it gets into contact with plant communities dominated by the native halophyte A. portulacoides. Our experiment shows that A. portulacoides adult plants are not able to outcompete S. den- siflora clumps coming from seeds in controlled greenhouse conditions. Our results in greenhouse controlled conditions corroborated our hypothesis based on field observations since the presence of adult plants of Atriplex did not prevented the establishment and growth of S. densiflora that competed suc- cessfully with the native halophyte as observed in the field.
ACKNOWLEDGEMENT
The author is grateful to assistance in the field (collect seed materials): David. We thank Jesús López and José María Car- ranza, Greenhouse general service (CITIUS), Facility of the University of Seville. This work was supported by Egyptian Government-ministry of higher education (cultural affairs and missions sector) PhD scholarship.
REFERENCES
[1] M. Williamson, "Biological Invasions", Chapman and
Hall, London, 1996.
[2] M. Vila`, and J. Weiner, "Are invasive plant species better competitors than native plant species? – evidence from pair-wise experiments", Oikos, 105, 229–238, 2004.
[3] J. Bakker, and S. D. Wilson, "Competitive abilities of in- troduced and native grasses", Plant. Ecol., 157, 119–127,
2001
[4] I. A. Ungar, "Are biotic factors significant in influencing the distribution of halophytes in saline habitats?", Bot. Rev., 64, 176-199, 1998.
[5] J. M., Levine, J. S. Brewer, and M. D. Bertness, "Nutrients, competition and plant zonation in a New England salt marsh", J. Ecol., 86, 285–292, 1998.
[6] J. M. Castillo, E. Mateos-Naranjo, F. J. Nieva, and E.
Figueroa, "Plant zonation at salt marshes of the endan- gered cordgrass Spartina maritima invaded by Spartina densiflora", Hydrobiologia, 614, 363–371, 2008.
[7] A. Bortolus, "The austral cordgrass Spartina densiflora Brong.: its taxonomy, biogeography and natural histo- ry", J. Biogeogr., 33, 158–168, 2006.
[8] V. Bouchard, and J. C. Lefeuvre, "Heterogeneity of Atri- plex portulacoides (L.) Aellen productivity in a macrotid- al salt marsh", Comptes Rendus de l’academie des Sci- ences serie III-Sciences de la Vie-Life Sciences 319, 1027-
1034, 1996.
[9] M. Mohamed, "Ecological studies on Atriplex portulacoides and its role in saltmarsh zonation", PhD Thesis. Univer- sity of East Anglia, Norwich, United Kingdom. 1998.
[10] C. Gaudet, and P. Keddy, "Competitive performance and species distribution in shoreline plant communities: a comparative approach", Ecology, 76, 280–291, 1995.
[11] E. Castellanos, M. Figueroa, and A. Davy, "Nucleation and facilitation in saltmarsh succession: interactions be- tween Spartina maritima and Arthrocnemum perenne", J. Ecol., 82, 239–248, 1994.
[12] H. K. Lichtenthaler, "Chlorophylls and carotenoids: Pig- ments of photosynthetic biomembranes", Methods in Enzymol., 148, 350–382, 1987.
[13] T. Ingestad, and A.B. Lund, "Theory and techniques for steady state mineral nutrition and growth of plants", Scan. J. For. Res., 1, 439—453, 1986.
[14] P. Kittelson, and M. Boyd, "Mechanisms of expansion for an introduced species of cordgrass, Spartina densiflo- ra, in Humboldt Bay, California", Estuaries, 20, 773-778,
1997.
[15] C. Dormann, R. Van der Wal, and J. Bakker, "Competi- tion and herbivory during salt marsh succession: the importance of forb growth strategy", J., Ecol., 88, 571-
583, 2000.
[16] A. Laffaille, P. Petillon, J. Parlier, E. Valery, L. Ysnel, F.
Radureau, A. E. Feunteun, and J. Lefeuvre, Does the
invasive plant Elymus athericus modify fish diet in tidal salt marshes? Est. Coast. Shelf Sci., 65, 739-746, 2005.
IJSER © 2014 http://www.ijser.org